




编辑/凯霞
人脑包含大约 1000 亿个神经元,每个神经元都有数千个突触连接。尽管单个神经元是神经系统的基本单位,但正是它们的突触连接模式使神经元能够为特定功能形成专门的神经环路,从而使大脑成为强大的计算设备。
近日,来自斯坦福大学生物系骆利群院士在《Science》发表综述文章《Architectures of neuronal circuits》。
该综述回顾了在不同大脑区域和动物物种中使用的常见环路模体(circuit motifs)和环路架构计划。
作者认为,了解突触连接的特定模式如何实现特定的神经计算,有助于弥合单个神经元生物学与整个大脑功能之间的巨大差距,使我们能够更好地理解行为的神经基础,并可能激发人工智能的新进展。

作者把神经元比喻为字母,大脑比喻为整篇文章,而微环路就是字母组成的单词,神经环路则是单词组成的句子。不同脑区使用的不同单词就是环路模体,而环路模体又进一步组成了复杂的神经环路架构。
常见的环路模体

图示:常见的电路图案。(来源:论文)
常见的环路模体有四种:前馈激活(Feedforward excitation)、前馈抑制和反馈抑制(Feedforward and feedback inhibition)、横向抑制(Lateral inhibition)、相互抑制(Mutual inhibition)。
前馈激活
信号从一个神经区域流向另一个神经区域的主要方式是前馈激活,即兴奋性神经元之间的一系列连接。
前馈激活最典型的例子之一是哺乳动物视觉系统,其中信号从光感受器流动 → 双极细胞 → 视网膜神经节细胞 → 外侧膝状核 (LGN) 中继神经元 → 第 4 层初级视觉皮层 (V1) 神经元 → V1 神经元其他层 → 较高皮层区域的神经元。哺乳动物视觉系统的前馈架构激发了用于图像识别和分类的「感知器」和「深度神经网络」的开发;深度神经网络在人工智能中也被用于解决远远超出图像分析的问题。
前馈抑制和反馈抑制
在前馈抑制中,抑制性神经元接收来自突触前兴奋性神经元的输入,抑制性和突触前兴奋性输入都汇聚到突触后神经元上。在反馈抑制中,抑制性神经元接收来自兴奋性神经元的输入并将其投射回兴奋性神经元,通常在其突触前末梢。
前馈抑制比反馈抑制作用更快,因为它到达突触后靶细胞仅在兴奋信号后有一个突触延迟,而反馈抑制有两个突触延迟。前馈抑制与输入强度成正比,而反馈抑制与输出强度成正比;两者都用于调节传入兴奋信号的持续时间和幅度。
横向抑制
横向抑制是一个广泛存在的环路模体。它通过放大并行路径之间的活动差异来选择要传播到下游环路的信息。横向抑制也用于其他感觉系统,其一般目的是使下游电路处理的行为学相关信息的表征更加清晰。
相互抑制
抑制性神经元之间的通信可以赋予环路有趣的特性。如果抑制性神经元 A 直接抑制抑制性神经元 B,则激活 A 将解除对 B 的目标神经元的抑制。如果 B 也抑制 A,则它们形成相互抑制图案。
相互抑制广泛用于表现出节律活动的回路,例如与运动有关的回路。一个典型的例子是甲壳动物的口胃神经节。
针对特定功能的特定环路架构
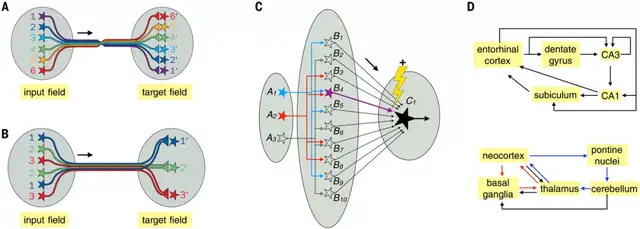
图示:特定功能的专门架构。(来源:论文)
连续地形图
连续地形图是表示神经系统信息的常见架构。相邻的输入神经元通过有序的轴突投影连接到相邻的目标神经元。一个主要的例子是视网膜移植术。
地形图提供了一种在处理的连续阶段组织信息的便捷方式,并且可以通过强大的发展机制来构建。它们具有多种计算优势。例如,「卷积神经网络」的设计借鉴了地形图,大大减少了调整人工神经网络所需的变量数量,从而加快了计算速度。
离散并行处理
离散并行处理允许通过离散信息通道并行表示和处理信号。一个典型的例子是脊椎动物嗅球和昆虫触角叶:表达相同气味受体的嗅觉受体神经元散乱分布在嗅球中,却将它们的轴突发送到同一个嗅小球上,从而提高了信噪比。
离散并行处理通常与连续地形图结合使用。与串行处理相比,并行处理降低了计算深度,从而降低了错误率,提高了处理速度。
维度扩展
在维数扩展架构中,来自相对少量输入神经元的信号发散到大量输出神经元上,允许输出神经元表示不同的输入组合。两个主要的例子是昆虫蘑菇体和脊椎动物小脑。
维度扩展的另一个例子是内嗅皮层 → 齿状回颗粒细胞 → CA3 锥体神经元回路。与蘑菇体和小脑皮层不同,这里还没有发现教学神经元。这可能是因为海马回路使用无监督学习,而小脑和蘑菇体回路实施类似于监督和强化学习的算法。
循环回路
神经系统充满了循环回路,其中神经元通常通过中间神经元连接回自身。这些循环回路在规模上是异质的,范围从特定神经区域内到跨越大脑的大部分区域。远程循环回路可能包含连续地形映射或离散并行处理架构。
偏倚输入—分离输出
调节神经元在神经系统中也有重要功能。小鼠中的病毒遗传追踪显示,中脑多巴胺、中缝背侧血清素和下丘脑神经肽甘丙肽系统都采用了群体水平的「偏倚输入—分离输出」架构。

图示:具有广泛投影的神经调节回路的输入-输出组织。(来源:论文)
进化和发展的观点
计算机电路是自上而下设计的产物,而复杂的神经元环路已经进化了数亿年。神经元回路也在发育过程中使用进化选择的遗传指令自组装,并通过经验进行微调。因此,现有的神经元环路结构很可能是在开发过程中很容易进化和组装的那些的选择。
孤立地观察神经元回路可能无法告诉我们哪些元素在功能上很重要。然而,观察通过趋同进化而被进化选择、扩大、缩小、消除或重复产生的东西,可以表明在功能研究中应该关注哪些元素。
神经元环路的进化
现存的双侧神经系统可能通过渐进的复杂性源自祖先。一些核心模体的普遍存在,例如前馈激活和前馈和反馈抑制,可能起源于具有中间神经元和中枢神经系统的动物,由于其实用性而在不同物种中保守,并在每个物种内的神经区域中传播。
昆虫和脊椎动物嗅觉系统的肾小球组织很可能是趋同进化的结果。视觉系统提供了许多基本特征的收敛进化的惊人例子,从视网膜到无脊椎动物和脊椎动物谱系中的运动检测算法。
大脑区域进化的复制和发散原则上应该使神经元回路模块化:复制单元内的丰富连接和单元之间的稀疏连接(与在许多人工神经网络中用作起始条件的全对全非模块化架构相反)。反过来,神经元回路的模块化特性可能会加速进化,因为不同的模块可以相互独立地进化。
神经环路的发展
进化主要通过在发育过程中修改参与环路布线的基因来对神经元环路施加影响。一个关键问题是,有限数量的基因如何构建具有大量突触连接和特定基序的神经系统。
细胞外信号及其细胞表面受体能够通过轴突和树突生长锥识别特定目标。这些分子是建立神经系统粗略组织的主要力量,还可以非常精确地指定某些环路和生物体中的突触连接。用有限数量的基因建立大量连接的特异性的一种策略,是使用相同蛋白质的不同表达水平来指定不同的连接。这种策略很容易用于构建连续的地形图。
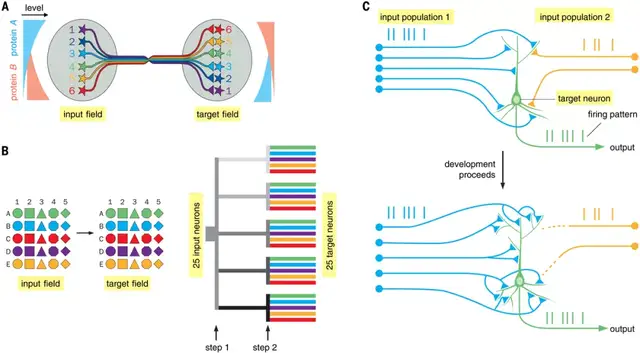
图示:连接神经元环路。(来源:论文)
有两种广泛的机制用于建立神经元回路的布线模式:分子线索硬连接神经系统和神经元活动,并体验微调连接。
展望
我们才刚刚开始深入了解在复杂神经系统中产生电路结构的进化和发育过程。例如,我们仍然不知道,神经元回路的布线过程和操作中的算法变化在多大程度上可以解释哺乳动物大脑复杂性的增加。
有意识地研究不同物种的关键环路架构中的字母是如何组合成单词,以及单词如何组合成句子,可能会产生有价值的见解。
使用单细胞转录组学对同源大脑区域的神经元类型组成进行比较研究,是第一步。接下来可以调查建立其连接模式并作为其功能操作基础的机制。整合对神经元环路的结构、功能、发育和进化的研究,将使人们能够更深入地了解超越单个神经元水平的神经系统组织。
此外,了解不同架构如何在单个神经系统中协作,也应该会激发新的人工神经网络,这些网络有朝一日可能会实现通用人工智能。
论文链接:https://www.science.org/doi/10.1126/science.abg7285
